EVOLUTION
![]()
![]()
![]()
![]()
![]()





Some fossil ascospores of undoubted Hypoxylon were described by Elsik (1990; see therein for a review of other possible fossil finds) among the 35 taxa of the fossil genus Hypoxylonites Elsik. Hypoxylonites is apparently an assemblage of fossil Xylariaceae as well as other unrelated fossil fungi, as suggested in Table 4 of Elsik (1990). According to Elsik (1990), the first specimens with a distinct Hypoxylonites morphology occur in the Middle Eocene. Kumar (1990) erected the fossil genus Hypoxylonsporites Kumar to accommodate two palynological species from the Miocene, and suggested its Hypoxylon affinity. Whether or not these first Hypoxylonites or Hypoxylonsporites are true Xylariaceae, the reliable fossil record of xylariaceous fungi is no earlier than the Middle Eocene. The extant fossil record of xylariaceous fungi is too recent to suggest the early evolutionary events which probably occurred in the Late Cretaceous or perhaps even earlier. We believe that the major diversification of the Xylariaceae is coincident with those of the angiosperms which had evidently occurred no later than the Late Cretaceous (Steward and Rothwell, 1993). Hence, for the present, we are compelled to base our evolutionary scheme on available data from extant xylariaceous fungi, and hope that our hypothesis can be tested through further findings of xylariaceous fossils and through the application of modern techniques.
Rogers (1979) stated that "the presence of cellular appendages on immature ascospores, in some cases persisting on the mature spore, indicates that the ancestors of the Xylariaceae had two-celled ascospores". As far as we know, there is no taxon in Hypoxylon possessing a cellular appendage on either mature or immature ascospores. However, in H. cohaerens (Rogers, 1972), H. multiforme (Rogers, 1967), and H. rubiginosum (Rogers, 1969), the immature ascospores at first contain two nuclei which result from a mitosis. One nucleus then degenerates as ascospores mature. However, in H. fuscum (Rogers, 1965), ascospores are uninucleate from formation to maturity. This indicates that, on the one hand, Hypoxylon is fairly recent for it no longer has two-celled ascospores, but, on the other hand, some taxa still retain the ancestral binucleate condition.
Xylariaceous anamorphs can be classified into six general forms based on their conidiogenesis, conidiogenous cell morphology and proliferation, and conidium morphology. These general forms of anamorphs ought to be considered relatively conservative because a certain form of anamorph is usually found across several genera. In order to look for the possible origin of Hypoxylon, it seems logical to consider those genera closely related to Hypoxylon--those genera bearing the Nodulisporium-like anamorph-and still having two-celled ascospores. Biscogniauxia appears to be an immediate relative of Hypoxylon where some taxa still have two-celled ascospores and a germ slit is present in the dark, larger cell. These two-celled Biscogniauxia were disposed in section Appendiculata Whalley & Laessøe by Whalley et al. (1990a). It is likely that section Appendiculata has retained some ancient characteristics from which we can infer the polarity of certain character states.
Table 1. Stromatal types of Biscogniauxia based on anatomical features. Names of stromatal types correspond to the section names of Biscogniauxia.
| Stromatal type | Representative taxa |
| Biscogniauxia | B. bartholomaei and B. nummularia |
| Scleraster | B. cinereolilacina, B. marginata, and B. repanda |
| Appendiculata | section Appendiculata Whalley & Laessøe (Whalley et al., 1990), the "Hypoxylon" nummularium complex of Miller (1961) except for B. nummularia, B. albosticta, B. atropunctata, and B. mediterranea and its varieties |
Figure 1. A-G. Stromatal types and ascospores in various groups of Biscogniauxia and Hypoxylon. A-E. Biscogniauxia. A. Appendiculata-type stroma with an appendaged ascospore. B. Appendiculata- type stroma with a nonappendaged ascospore. C. A hypothetical stromatal type. D. Scleraster- type stroma. E. Biscogniauxia-type stroma. F and G. Hypoxylon. F. Stromatal type found in Hypoxylon section Annulata with the dehiscent ascospore perispore bearing a thickening. G. Stromatal type found in Hypoxylon section Hypoxylon with the dehiscent ascospore perispore bearing coil-like ornamentation. a: host tissue; b: outermost stromatal layer; c: waxy stromatal tissue; d: woody stromatal tissue; e: carbonaceous stromatal tissue.
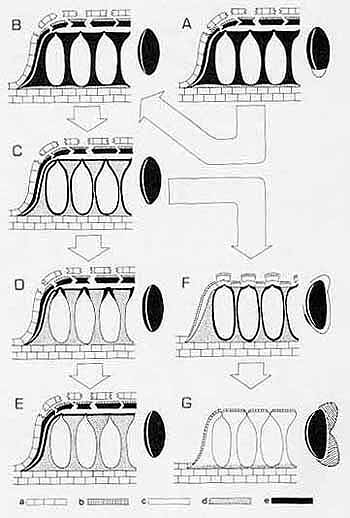
Stromata of Biscogniauxia are mainly composed of two types of stromatal tissues: black, carbonaceous stromatal tissue and brown, woody stromatal tissue. As shown in Fig. 1 and Table 1, three stromatal types can be recognized in Biscogniauxia. All of these stromatal types have a layer of carbonaceous tissue forming a crust on the surface which probably represents the lower part of the ectostromata from which the upper part of ectostromata has sloughed off. Based on the presence or absence of the extra carbonaceous stromatal tissue, and, if present, the distribution and the quantity of this extra carbonaceous stromatal tissue, these three stromatal types can be discerned. Biscogniauxia-type stromata (Fig. 1E) do not have extra carbonaceous stromatal tissue. Appendiculata-type stromata (Fig. 1A & 1B) have extra carbonaceous stromatal tissue occupying the interperithecial spaces. Scleraster-type stromata (Fig. 1D) have extra carbonaceous stromatal tissue much reduced to only encase each individual ostiole.
All known taxa of Biscogniauxia with two-celled ascospores also possess the Appendiculata-type stromata. It is probable that the Appendiculata-type stromata were possessed by the ancestral taxa of Biscogniauxia. Other Biscogniauxia that have the Appendiculata-type stromata, but have one-celled ascospores (Fig. 1B), may have been derived from two-celled Biscogniauxia by loss of the smaller hyaline cell of ascospores. They may have had the general look of extant taxa like B. albosticta Ellis & Morg., B. atropunctata (Schwein.: Fr.) Pouzar, B. mediterranea (De Not.) Kuntze and its varieties, and most taxa referred to the "Hypoxylon" nummularium complex by Miller (1961) except for B. nummularia. The other two types of stromata might have arisen by gradual reduction of the amount of the extra carbonaceous stromatal tissue surrounding the ostiolar regions only, as shown in the Scleraster-type, and eventually losing it, as shown in the Biscogniauxia-type. During this evolutionary process, before the carbonaceous stromatal tissue was much reduced, there might have been an intermediate type of stroma, possessing a carbonaceous stromatal tissue discretely enclosing each perithecium (Fig. 1C). Such a stroma is known in the curious genus, Induratia Samuels, E. Müller, & Petrini (Samuels et al., 1987). If Hypoxylon tormentosum Ces., which is only known from Sarawak, Malaysia, is eventually proven to be a Biscogniauxia, its stromata seem to be of this intermediate type (also see the entry tormentosum in the LIST). This particular type of stroma is universally found in taxa of Hypoxylon section Annulata (Figs. 1F). Like taxa of Biscogniauxia, most taxa of Hypoxylon section Annulata have Periconiella-like and Nodulisporium-like conidiogenous structures, and only a few taxa have Virgariella-like conidiogenous structures. Although Hypoxylon section Annulata does not have bipartite stromata as does Biscogniauxia, the mechanism for forming the ostiolar discs by pushing away the overlying stromatal tissue in the former group is similar to the separating of the ectostromata from the entostromata in the latter group. A layer of whitish palisade tissue is involved in separating the two kinds of stromatal tissues in both cases. Ostiolar discs in Hypoxylon section Annulata can be viewed as restricted ectostromata that are sloughed off, i.e., as the last vestiges of a bipartite stroma. It is thus probable that one of the evolutionary trends leading from Biscogniauxia to Hypoxylon section Annulata is to have a progressive fusing of the ectostromata with the entostromata except for the areas surrounding the ostioles. The stromata of some temperate Hypoxylon--H. cohaerens, H. multiforme and their respective varieties, and H. nothofagi--probably represent the final steps in the evolution of this character where the ostiolar discs are nearly or entirely undetectable. The fusion of ectostromata and entostromata may have accompanied the evolution of stromata that are superficial on substrates. A separable ectostroma functioning in dehiscence of the overlying host substrates does not seem necessary in Hypoxylon section Annulata as it is in Biscogniauxia. Achieving a superficial position for the stromata in Hypoxylon could also account for the alteration in the morphology of its conidiogenous structures, mainly with the reduction of the main stipes. Anamorphs of both Biscogniauxia and Hypoxylon usually grow on the surface of developing stromata. The conidiogenous structures of Biscogniauxia, when found on the natural substrates, are uniformly Periconiella-like; those of Hypoxylon section Annulata range from Periconiella-like in a few taxa, Nodulisporium-like in the great majority, to Virgariella-like in a few taxa; those of Hypoxylon section Hypoxylon range from Periconiella-like in a few taxa, Nodulisporium-like and Virgariella-like in the great majority, to Sporothrix-like in a few taxa. It can be hypothesized that-after stromata had evolved from being within the host substrates, as seen in Biscogniauxia, to superficial on the host substrates, as seen in most Hypoxylon--a long, stout main stipe was not necessary for efficient discharge of conidia.
Some morphological changes also occurred during the evolutionary development of Hypoxylon section Annulata. The carbonaceous stromatal crust became more waxy and came to contain KOH-extractable pigments which are either olivaceous or purplish; the stromatal surface became colorful, probably vinaceous or with vinaceous shades. Stromatal pigments probably were favored as stromata became superficial as contrasted with development within bark. Pigments are probably involved as photosensitive elements in stromatal physiology. The asymmetrical ascospores became dominant and the perispore became dehiscent. It is noteworthy that most extant taxa of Hypoxylon section Annulata have the germ slit on the convex side of ascospores. Asymmetrical ascospores with the germ slit on the convex side seem to provide an easy and quick way for germination when there is sufficient moisture. This device may have been efficient for Hypoxylon section Annulata to have adapted to the seasonally dry areas into which their angiospermous hosts had rapidly invaded. However, the easily opened ascospore wall is apparently architecturally weak because a structural discontinuity exists at the germ slit. Therefore, the outermost hyaline wall layer evolved into a separable layer, the perispore, which is equipped with a thickening positioned at ca. 1/3 ascospore length on the same side as the germ slit (see section Annulata). The thickening apparently exerts a transverse constraint to counteract a break-up along the germ slit.
Three curious taxa in Hypoxylon section Annulata--H. leptascum, H. leptascum var. macrosporum, and H. urceolatum--appear to form a coherent group. These fungi have pale brown ascospores which have an indehiscent perispore and have a very short germ slit that extends from one end of the ascospore. The hypothesis that the short germ slit extending from only one end of an ascospore may have been present in some ancestors of Hypoxylon section Annulata provides a logical explanation for the position of the thickening on the perispore in other taxa of Hypoxylon section Annulata. Some early members of Hypoxylon section Annulata might have had a short germ slit extending only from one end. Their subsequent successors might have evolved to have asymmetrical ascospores and a dehiscent perispore, and to have a thickening on the perispore at the position opposing the germ slit. Ascospores of this morphology can be found in some extant taxa like H. bovei, H. multiforme, H. multiforme var. alaskense, and H. cohaerens var. microsporum. These taxa have a less than spore-length germ slit, and, in a large proportion of ascospores, the position of the germ slit tends to correspond to that of the thickening on the perispore at ca. 1/3 ascospore length. After having evolved the thickening on the perispore, the elongation of the germ slit to spore-length took place to complete the commonest form of the ascospores in Hypoxylon section Annulata. This may explain why extant taxa of Hypoxylon section Annulata with the spore-length germ slit still exclusively retain the perispore thickening at that particular position.
Hypoxylon section Hypoxylon differs from Hypoxylon section Annulata in lacking a carbonaceous stromatal tissue surrounding each perithecium (Fig. 1G), in having coil-like ornamentation rather than a thickening on the perispore, and in not having ostiolar discs. Putative evidence that Hypoxylon section Hypoxylon was derived from Hypoxylon section Annulata primarily relates to the fact that a few collections of H. subrutilum from Venezuela show both the thickening and coil-like ornamentation on the same perispore. This type of ascospore is also believed to have been present in those--probably extinct--transitional forms which were evolving away from Hypoxylon section Annulata and towards Hypoxylon section Hypoxylon. During this transition, some major modifications can be inferred by comparing the extant taxa in both sections. With the increasing softness or with the delay of the hardening of the overall stromatal tissue, the requirement for perithecia to make their way through the surrounding stromatal tissue with the aid of a layer of carbonaceous stromatal tissue had gradually been minimized. No carbonaceous stromatal tissue surrounding individual perithecia is found in the extant Hypoxylon section Hypoxylon. Perithecia have become directly embedded in either woody or, in most taxa, waxy stromatal tissues. The thickening on the perispore in Hypoxylon section Annulata had gradually been replaced by the coil-like ornamentation in Hypoxylon section Hypoxylon. Ostioles gradually retreated from a position that is higher than the stromatal surface in section Annulata to one that is slightly higher or, in most extant taxa, lower than the stromatal surface in section Hypoxylon. The production of orange stromatal pigments appears to be a novelty originating in Hypoxylon section Hypoxylon, as it is not found in Hypoxylon section Annulata. The orange stromatal pigments may not have been present in the earlier Hypoxylon section Hypoxylon. In summary, the early Hypoxylon section Hypoxylon may have had the following characteristics: stromata with dull-colored waxy stromatal tissue which contains either olivaceous or purplish KOH-extractable pigments; perithecia directly embedded in the waxy or woody stromatal tissue; ostioles slightly higher than, or at the same level as, the stromatal surface; ascospores asymmetrical, dark-colored, narrow-ended, with the spore-length germ slit situated on the convex side; conidiogenous structure either Periconiella-like or Nodulisporium-like.
Some taxa in both sections of Hypoxylon subsequently lost the dehiscing perispore, lost the inequality of ascospores, and/or shortened the germ slit. It is possible that these processes were accompanied by their angiospermous hosts adapting to areas with more mesic environmental conditions so that tolerance to desiccation was no longer a major determining factor in selecting certain morphologies for ascospores. Another feature that probably came after Hypoxylon section Hypoxylon had evolved is the loss of ascal apical ring. The ringless ascus is not known in Biscogniauxia or Hypoxylon section Annulata, but is seen in several taxa of Hypoxylon section Hypoxylon and Rhopalostroma among the Hypoxyloideae. These taxa of section Hypoxylon (see NOTES on H. commutatum) do not appear to be closely related among themselves or to taxa of Rhopalostroma. This raises the probability that the loss of the ascal apical ring occurred more than once. This in turn suggests that these fungi may have become less dependent on air to disperse the sexual propagules. Other agents such as water or insects may have been employed to spread the sexual propagules.
Several genera that are undoubtedly closely related to Hypoxylon section Hypoxylon may have evolved from the latter and may be considered to be advanced genera in the Hypoxyloideae. Rhopalostroma, lacking the ascal apical ring as just mentioned above, adopted an erect stromatal habit. Other advanced characteristics found in this genus include less inequilateral ascospores and indehiscent perispores. This genus may have arisen from the H. placentiforme line which comprises taxa with massive stromata, including H. placentiforme, H. begae, H. kretzschmarioides, H. tortisporum, H. symphyon, H. nicaraguense, H. polyporum, and H. sclerophaeum. Daldinia, which is characterized by having concentric rings in the stromata, is another genus that could have arisen from the H. placentiforme line. It has been shown that ascospores of Daldinia concentrica have a dehiscent perispore with coil-like ornamentation (Beckett, 1976a). Furthermore, a layer of waxy stromatal tissue is formed immediately beneath the surface; it has a Nodulisporium-like conidiogenous structure. All of these characteristics, except for the concentric rings, also exist in H. placentiforme. Entonaema is characterized by having orange granules containing orange pigments and having a gelatinous layer lining a cavity. The gelatinous matrix is known to have a water-holding capacity. A Nodulisporium-like conidiogenous structure has been reported by Rogers (1982). This genus may have evolved from taxa of Hypoxylon section Hypoxylon that have orange granules. Besides the orange granules and the gelatinous matrix, taxa of Entonaema also have an indehiscent perispore and nearly equilateral ascospores.